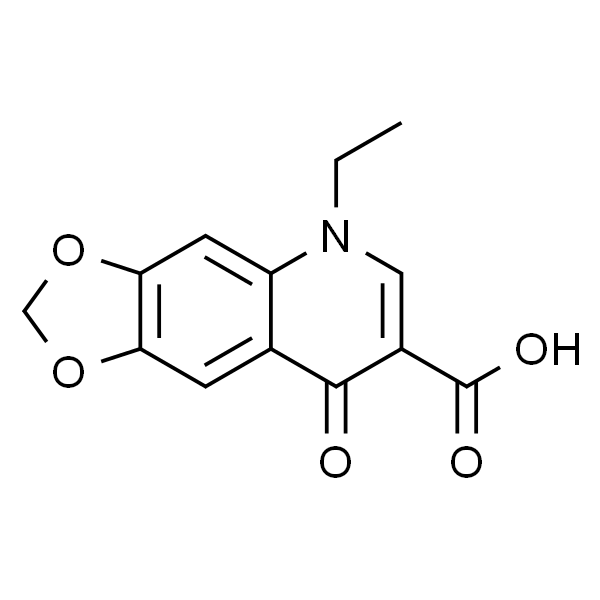
恶喹酸
货号:
IO0890
品牌:
Jinpan
暂无详情
产品简介
| EC | EINECS 238-750-8 |
| MDL | MFCD00056775 |
| InChIKey | KYGZCKSPAKDVKC-UHFFFAOYSA-N |
| InChI | InChI=1S/C13H11NO5/c1-2-14-5-8(13(16)17)12(15)7-3-10-11(4-9(7)14)19-6-18-10/h3-5H,2,6H2,1H3,(H,16,17) |
| PubChem CID | 4628 |
| 别名 | 奥索利酸; NSC-110364 |
| 英文名称 | Oxolinic Acid |
| CAS | 14698-29-4 |
| 分子式 | C13H11NO5 |
| 分子量 | 261.23 |
| 储存条件 | 2-8℃ |
| 纯度 | ≥98% |
| 单位 | 瓶 |
| SMILES | O=C(C1=CN(CC)C2=C(C=C3C(OCO3)=C2)C1=O)O |
| 靶点 | DNA/RNA Synthesis |
| 规格 | 500mg 1g |
Oxolinic acid抑制DNA促旋酶和DNA合成。它还是一种多巴胺再摄取的抑制剂。